Behavioral Neuroscience
Summers
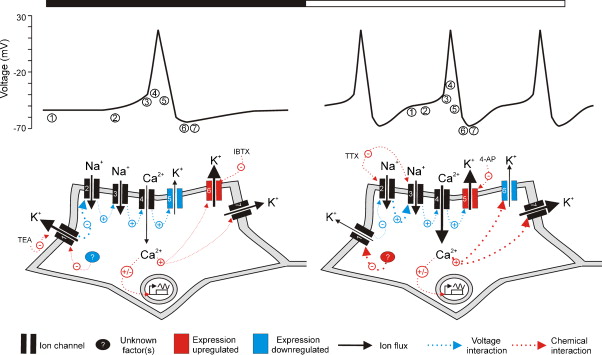
Sensory input for Rhythmicity
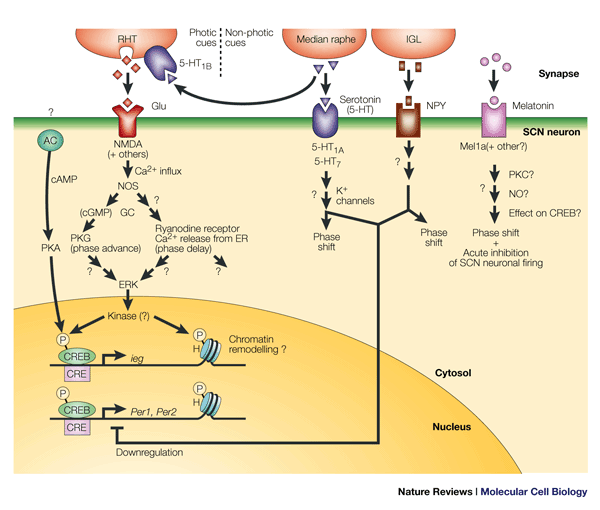
Afferent path to the SCN
Figures of Rhythmicity
Retina-RGC-SCN
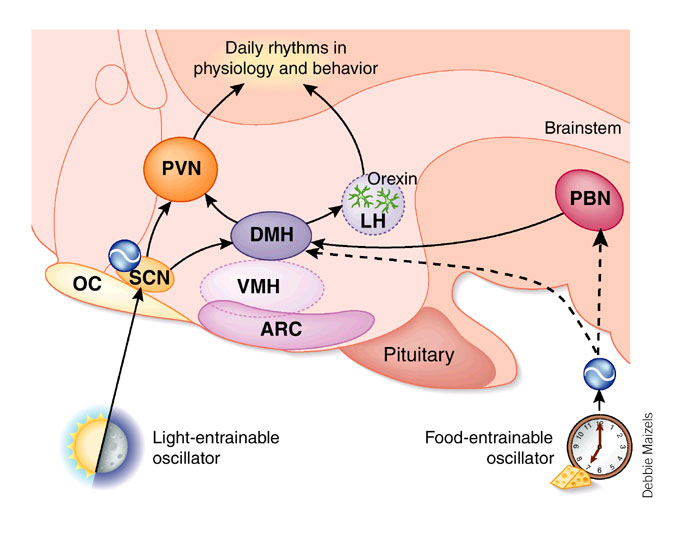
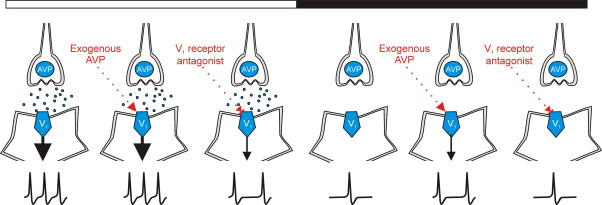
Efferent SCN output
Integration of Rhythms into Behavior
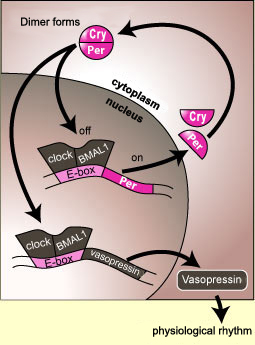
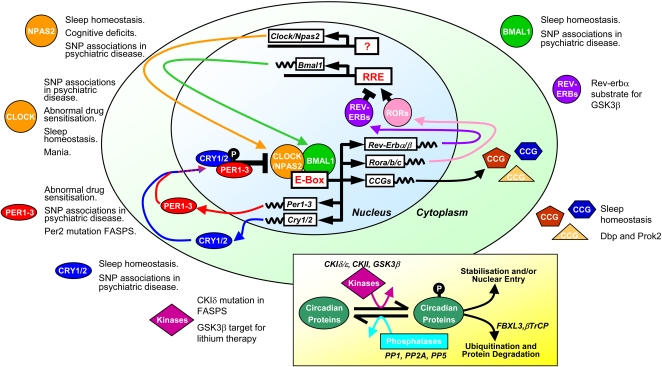
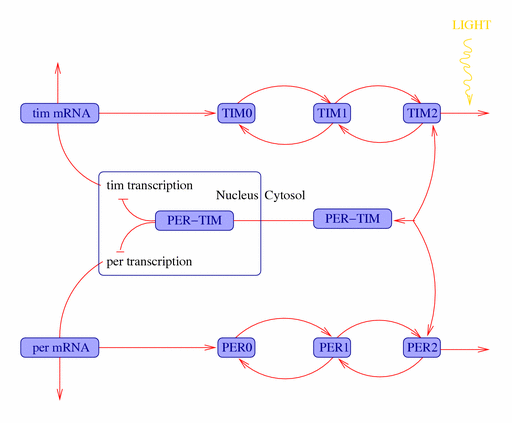
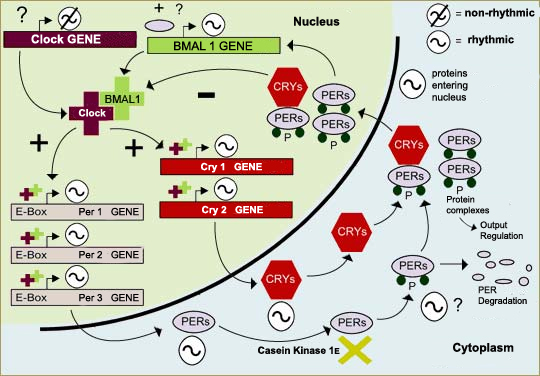
Molecular SCN
end Acronyms/Abbreviations Syllabus
VIP
AVP
GABA
5-HT
 the Brain from Top to Bottom
the Brain from Top to Bottom